Emergence of Resistance to Fluoroquinolones and Third-Generation Cephalosporins in Salmonella Typhi in Lahore, Pakistan

 ,
,  ,
,  , ,
, , 
 , and
, and 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Isolates
2.2. DNA Extraction
2.3. Whole Genome Nucleotide Sequencing
2.4. Genome Assembly and Annotation
2.5. Genotyping and Antimicrobial Gene Prediction
2.6. Phylogenetic Analysis
2.7. Role of the Funding Source
2.8. Ethics Statement
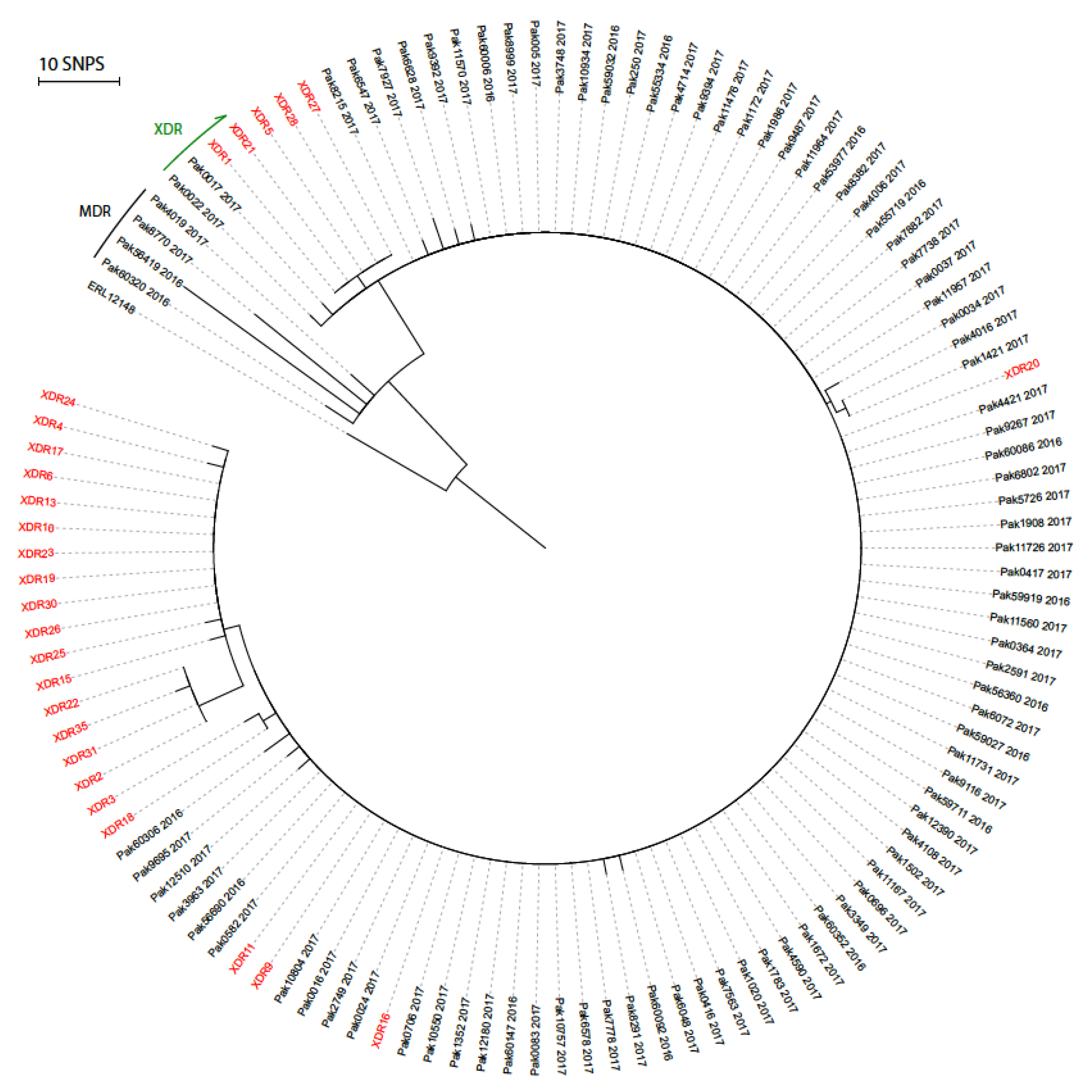
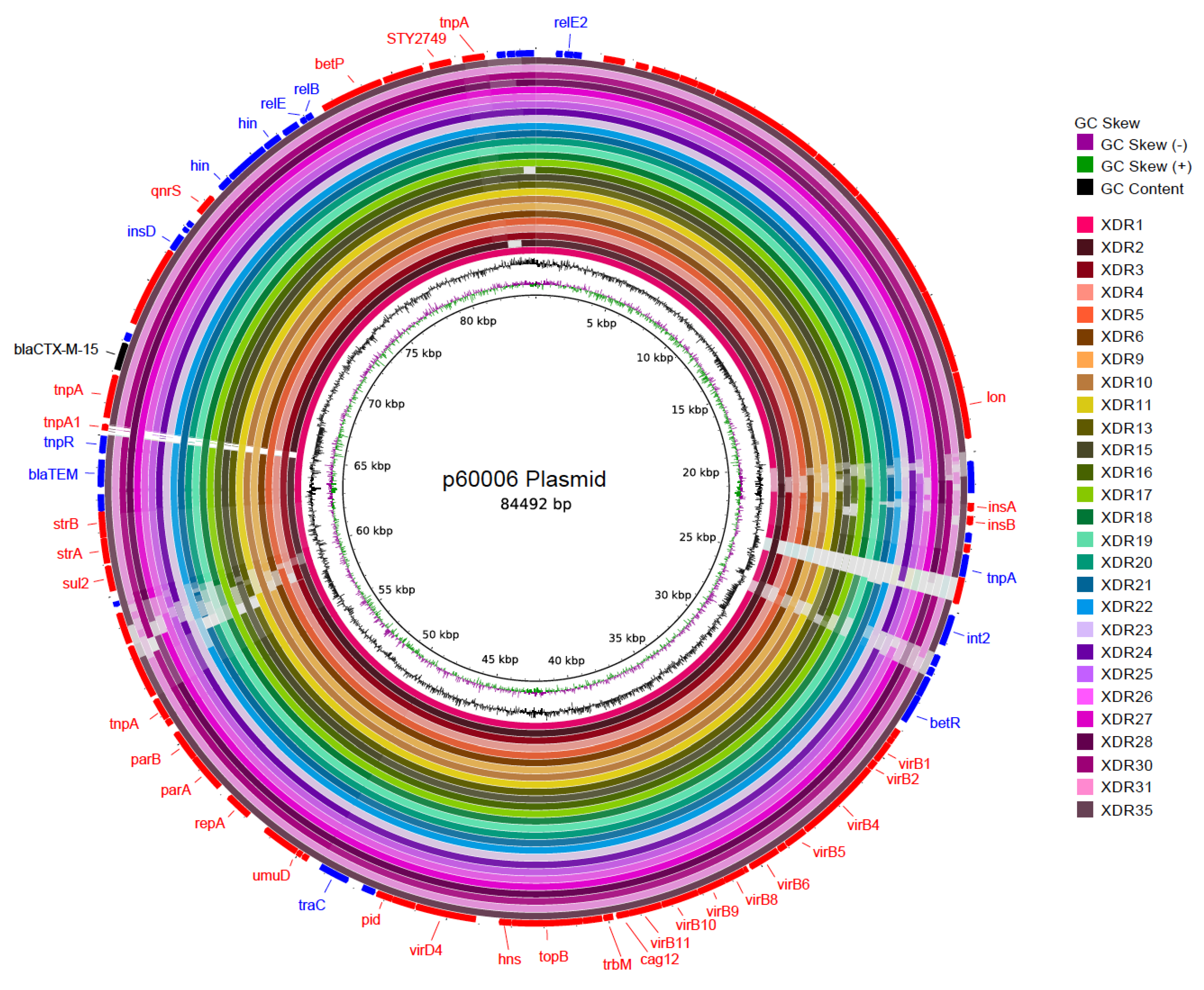
3. Results
3.1. Phenotypic Findings
3.2. Genotypic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buckle, G.C.; Walker, C.L.; Black, R.E. Typhoid fever and paratyphoid fever: Systematic review to estimate global morbidity and mortality for 2010. J. Glob. Health 2012, 2, 010401. [Google Scholar] [CrossRef] [PubMed]
- Mogasale, V.; Maskery, B.; Ochiai, R.L.; Lee, J.S.; Mogasale, V.V.; Ramani, E.; Kim, Y.E.; Park, J.K.; Wierzba, T.F. Burden of typhoid fever in low-income and middle-income countries: A systematic, literature-based update with risk-factor adjustment. Lancet Glob. Health 2014, 2, e570–e580. [Google Scholar] [CrossRef] [Green Version]
- Wain, J.; Kidgell, C. The emergence of multidrug resistance to antimicrobial agents for the treatment of typhoid fever. Tans. R. Soc. Trop. Med. Hyg. 2004, 98, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Bhutta, Z.A. Current concepts in the diagnosis and treatment of typhoid fever. Br. Med. J. 2006, 333, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klemm, E.J.; Shakoor, S.; Page, A.J.; Qamar, F.N.; Judge, K.; Saeed, D.K.; Wong, V.K.; Dallman, T.J.; Nair, S.; Baker, S.; et al. Emergence of an Extensively Drug-Resistant Salmonella enterica Serovar Typhi Clone Harboring a Promiscuous Plasmid Encoding Resistance to Fluoroquinolones and Third-Generation Cephalosporins. MBio 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Institute of Health (NIH) Islamabad. Weekly Field Epidemiology Report. 2020, pp. 1–8. Available online: https://www.nih.org.pk/wp-content/uploads/2020/07/26-FELTP-Pakistan-Weekly-Epidemiological-Report-June-21-27-2020.pdf (accessed on 23 July 2020).
- Muhammad, S.; Muhammad Hidayat, R.; Farhan, R.; Muhammad, S.; Muhammad Atif, N.; Ambreen Anwar, I.; Saba, T.; Afreenish, A.; Aamer, I.; Mohsin, K. Extended-spectrum beta-lactamases producing extensively drug-resistant Salmonella Typhi in Punjab, Pakistan. J. Infect. Dev. Ctries. 2020, 14, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Saeed, N.; Usman, M.; Khan, E.A. An Overview of Extensively Drug-resistant Salmonella Typhi from a Tertiary Care Hospital in Pakistan. Cureus 2019, 11, e5663. [Google Scholar] [CrossRef] [Green Version]
- Sah, R.; Donovan, S.; Seth-Smith, H.M.; Bloemberg, G.; Wüthrich, D.; Stephan, R.; Kataria, S.; Kumar, M.; Singla, S.; Deswal, V. A novel lineage of ceftriaxone-resistant Salmonella Typhi from India that is closely related to XDR S. Typhi found in Pakistan. Clin. Infect. Dis. 2020, 71, 1327–1330. [Google Scholar] [CrossRef]
- Watkins, L.K.F.; Winstead, A.; Appiah, G.D.; Friedman, C.R.; Medalla, F.; Hughes, M.J.; Birhane, M.G.; Schneider, Z.D.; Marcenac, P.; Hanna, S.S. Update on Extensively Drug-Resistant Salmonella Serotype Typhi Infections Among Travelers to or from Pakistan and Report of Ceftriaxone-Resistant Salmonella Serotype Typhi Infections Among Travelers to Iraq—United States, 2018–2019. Morb. Mortal. Weekly Rep. 2020, 69, 618. [Google Scholar] [CrossRef]
- Haqqi, A.; Khurram, M.; Din, M.S.U.; Aftab, M.N.; Ali, M.; Ahmed, H.; Afzal, M.S. COVID-19 and Salmonella Typhi co-epidemics in Pakistan: A real problem. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Katiyar, A.; Sharma, P.; Dahiya, S.; Singh, H.; Kapil, A.; Kaur, P. Genomic profiling of antimicrobial resistance genes in clinical isolates of Salmonella Typhi from patients infected with Typhoid fever in India. Sci. Rep. 2020, 10, 8299. [Google Scholar] [CrossRef] [PubMed]
- The COVID-19 Genomics UK (COG-UK) consortium. An integrated national scale SARS-CoV-2 genomic surveillance network. Lancet Microbe 2020, 1, e99–e100. [Google Scholar] [CrossRef]
- Liu, P.-Y.; Wang, K.-C.; Hong, Y.-P.; Chen, B.-H.; Shi, Z.-Y.; Chiou, C.-S. The first imported case of extensively drug-resistant Salmonella enterica serotype Typhi infection in Taiwan and the antimicrobial therapy. J. Microbiol. Immunol. Infect. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.; Al Rawahi, H.; Patel, S.; Yau, Y.; Eshaghi, A.; Zittermann, S.; Tattum, L.; Morris, S.K. The first Canadian pediatric case of extensively drug-resistant Salmonella Typhi originating from an outbreak in Pakistan and its implication for empiric antimicrobial choices. IDCases 2019, 15, e00492. [Google Scholar] [CrossRef]
- Engsbro, A.L.; Riis Jespersen, H.S.; Goldschmidt, M.I.; Mollerup, S.; Worning, P.; Pedersen, M.S.; Westh, H.; Schneider, U.V. Ceftriaxone-resistant Salmonella enterica serotype Typhi in a pregnant traveller returning from Karachi, Pakistan to Denmark, 2019. Euro. Surveill. 2019, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Song, L.; Breitwieser, F.P.; Salzberg, S.L. Centrifuge: Rapid and sensitive classification of metagenomic sequences. Genome Res. 2016, 26, 1721–1729. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Powers, C.M.; Grieger, K.D.; Hendren, C.O.; Meacham, C.A.; Gurevich, G.; Lassiter, M.G.; Money, E.S.; Lloyd, J.M.; Beaulieu, S.M. A web-based tool to engage stakeholders in informing research planning for future decisions on emerging materials. Sci. Total Environ. 2014, 470–471, 660–668. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.-F.; Mohamed, K.; Group TAS; Achtman, M. The user’s guide to comparative genomics with EnteroBase. Three case studies: Micro-clades within Salmonella enterica serovar Agama, ancient and modern populations of Yersinia pestis, and core genomic diversity of all Escherichia. BioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Wong, V.K.; Baker, S.; Connor, T.R.; Pickard, D.; Page, A.J.; Dave, J.; Murphy, N.; Holliman, R.; Sefton, A.; Millar, M.; et al. An extended genotyping framework for Salmonella enterica serovar Typhi, the cause of human typhoid. Nat. Commun. 2016, 7, 12827. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.; Mather, A.E.; Sanchez-Buso, L.; Page, A.J.; Parkhill, J.; Keane, J.A.; Harris, S.R. ARIBA: Rapid antimicrobial resistance genotyping directly from sequencing reads. Microb. Genom. 2017, 3, e000131. [Google Scholar] [CrossRef] [PubMed]
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef] [Green Version]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid phylogenetic analysis of large samples of recombinant bacterial whole genome sequences using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Alikhan, N.-F.; Petty, N.K.; Zakour, N.L.B.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [Green Version]
- Hameed, S.; Izhar, M.; Basheer, A.; Lal, C.; Rishi, S.; Basit, A. An Update on Isolation of Extensively Drug Resistant (XDR) Salmonella enterica from Blood Cultures in a Tertiary Care Centre. Proc. S.Z.P.G.M.I. 2019, 33, 5. [Google Scholar]
- Kidgell, C.; Reichard, U.; Wain, J.; Linz, B.; Torpdahl, M.; Dougan, G.; Achtman, M. Salmonella typhi, the causative agent of typhoid fever, is approximately 50,000 years old. Infect. Genet. Evol 2002, 2, 39–45. [Google Scholar] [CrossRef]
- Hafeez, M.; Saleem, Z.; Bukhari, N.A.; Hussain, K.; Shamim, R.; Hussain, A.; Bukhari, N.I. Off-label antibiotic use in a specialized children care hospital in Punjab, Pakistan: Findings and implications. J. Infect. Dev. Ctries. 2020, 14, 540–544. [Google Scholar] [CrossRef]
- Hannan, A.; Yasir, M.; Tahir, N.A.; Izhar, M.; Arshad, M.; Arshad, M.U. Carbapenemase intrudes K. pneumoniae in local clinical isolates. Pak. J. Path. 2011, 22, 5. [Google Scholar]
- Hooda, Y.; Tanmoy, A.M.; Sajib, M.S.I.; Saha, S. Mass azithromycin administration: Considerations in an increasingly resistant world. BMJ Global Health 2020, 5, e002446. [Google Scholar] [CrossRef] [PubMed]
- Malhotra-Kumar, S.; Lammens, C.; Coenen, S.; Van Herck, K.; Goossens, H. Effect of azithromycin and clarithromycin therapy on pharyngeal carriage of macrolide-resistant streptococci in healthy volunteers: A randomised, double-blind, placebo-controlled study. Lancet 2007, 369, 482–490. [Google Scholar] [CrossRef]
- Hooda, Y.; Sajib, M.S.; Rahman, H.; Luby, S.P.; Bondy-Denomy, J.; Santosham, M.; Andrews, J.R.; Saha, S.K.; Saha, S. Molecular mechanism of azithromycin resistance among typhoidal Salmonella strains in Bangladesh identified through passive pediatric surveillance. PLoS Neglect. Trop. Dis. 2019, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Satti, L.; Hanif, F.; Zehra, N.M.; Nadeem, S.; Bangash, T.M.; Peter, A. Typhoidal Salmonella strains in Pakistan: An impending threat of extensively drug-resistant Salmonella Typhi. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 2145–2149. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Ashton, P.; Doumith, M.; Connell, S.; Painset, A.; Mwaigwisya, S.; Langridge, G.; de Pinna, E.; Godbole, G.; Day, M. WGS for surveillance of antimicrobial resistance: A pilot study to detect the prevalence and mechanism of resistance to azithromycin in a UK population of non-typhoidal Salmonella. J. Antimicrob. Chemother. 2016, 71, 3400–3408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosholder, A.D.; Mathew, J.; Alexander, J.J.; Smith, H.; Nambiar, S. Cardiovascular risks with azithromycin and other antibacterial drugs. N. Engl. J. Med. 2013, 368, 1665–1668. [Google Scholar] [CrossRef]
- Naz, F.; Yousafzai, M.; Khaliq, A.; Karim, S.; Memon, H.; Junejo, A.; Baig, I.; Rehman, N.; Bhurgry, S.; Afroz, H.; et al. Adverse events following immunization with typhoid conjugate vaccine in an outbreak setting in Hyderabad, Pakistan. Vaccine 2020, 38. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasheed, F.; Saeed, M.; Alikhan, N.-F.; Baker, D.; Khurshid, M.; Ainsworth, E.V.; Turner, A.K.; Imran, A.A.; Rasool, M.H.; Saqalein, M.; et al. Emergence of Resistance to Fluoroquinolones and Third-Generation Cephalosporins in Salmonella Typhi in Lahore, Pakistan. Microorganisms 2020, 8, 1336. https://doi.org/10.3390/microorganisms8091336
Rasheed F, Saeed M, Alikhan N-F, Baker D, Khurshid M, Ainsworth EV, Turner AK, Imran AA, Rasool MH, Saqalein M, et al. Emergence of Resistance to Fluoroquinolones and Third-Generation Cephalosporins in Salmonella Typhi in Lahore, Pakistan. Microorganisms. 2020; 8(9):1336. https://doi.org/10.3390/microorganisms8091336
Chicago/Turabian StyleRasheed, Farhan, Muhammad Saeed, Nabil-Fareed Alikhan, David Baker, Mohsin Khurshid, Emma V. Ainsworth, A. Keith Turner, Ambereen Anwar Imran, Muhammad Hidayat Rasool, Muhammad Saqalein, and et al. 2020. "Emergence of Resistance to Fluoroquinolones and Third-Generation Cephalosporins in Salmonella Typhi in Lahore, Pakistan" Microorganisms 8, no. 9: 1336. https://doi.org/10.3390/microorganisms8091336
APA StyleRasheed, F., Saeed, M., Alikhan, N.-F., Baker, D., Khurshid, M., Ainsworth, E. V., Turner, A. K., Imran, A. A., Rasool, M. H., Saqalein, M., Nisar, M. A., Fayyaz ur Rehman, M., Wain, J., Yasir, M., Langridge, G. C., & Ikram, A. (2020). Emergence of Resistance to Fluoroquinolones and Third-Generation Cephalosporins in Salmonella Typhi in Lahore, Pakistan. Microorganisms, 8(9), 1336. https://doi.org/10.3390/microorganisms8091336