The Effect of Sea Buckthorn (Hippophae rhamnoides L.) Seed Oil on UV-Induced Changes in Lipid Metabolism of Human Skin Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Examination of Sea Buckthorn Seed Oil Composition
2.1.1. Determination of Squalene Levels
2.1.2. Phytosterols Profile
2.1.3. Determination of β-Carotene Levels
2.2. Cell Culture
2.3. Examination of Pro-Oxidative Activity
2.3.1. Determination of ROS Generation
2.3.2. Determination of Pro-Oxidants Enzyme Activities
2.4. Examination of Antioxidant Defence System
2.4.1. Determination of Antioxidant Enzymes Activity
2.4.2. Determination of Non-Enzymatic Antioxidant Levels
2.4.3. Determination of Protein Expression
2.5. Examination of Lipid Metabolism
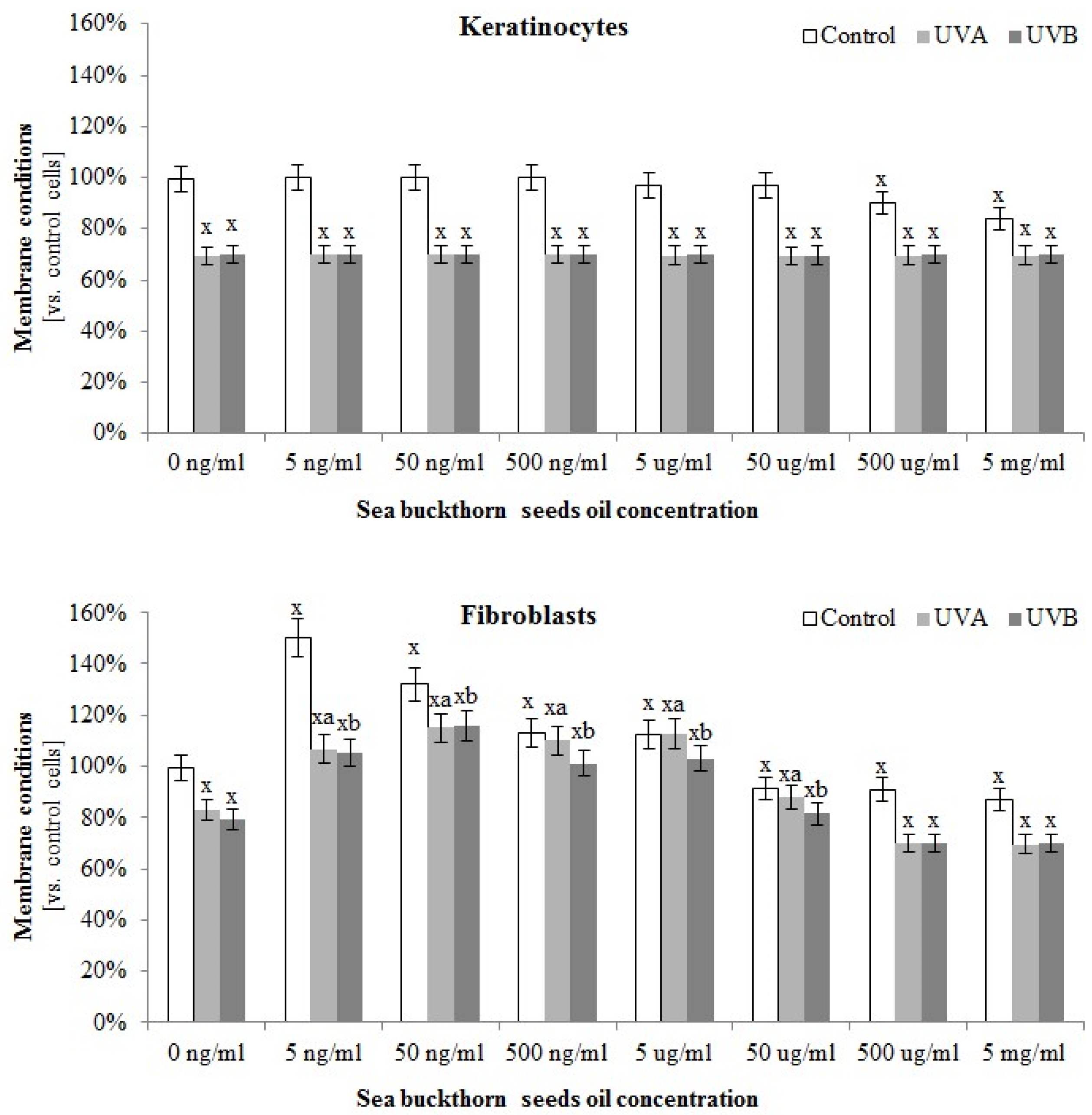
2.5.1. Determination of Cellular Membrane Integrity by LDH Test
2.5.2. Determination of Fatty Acids Levels
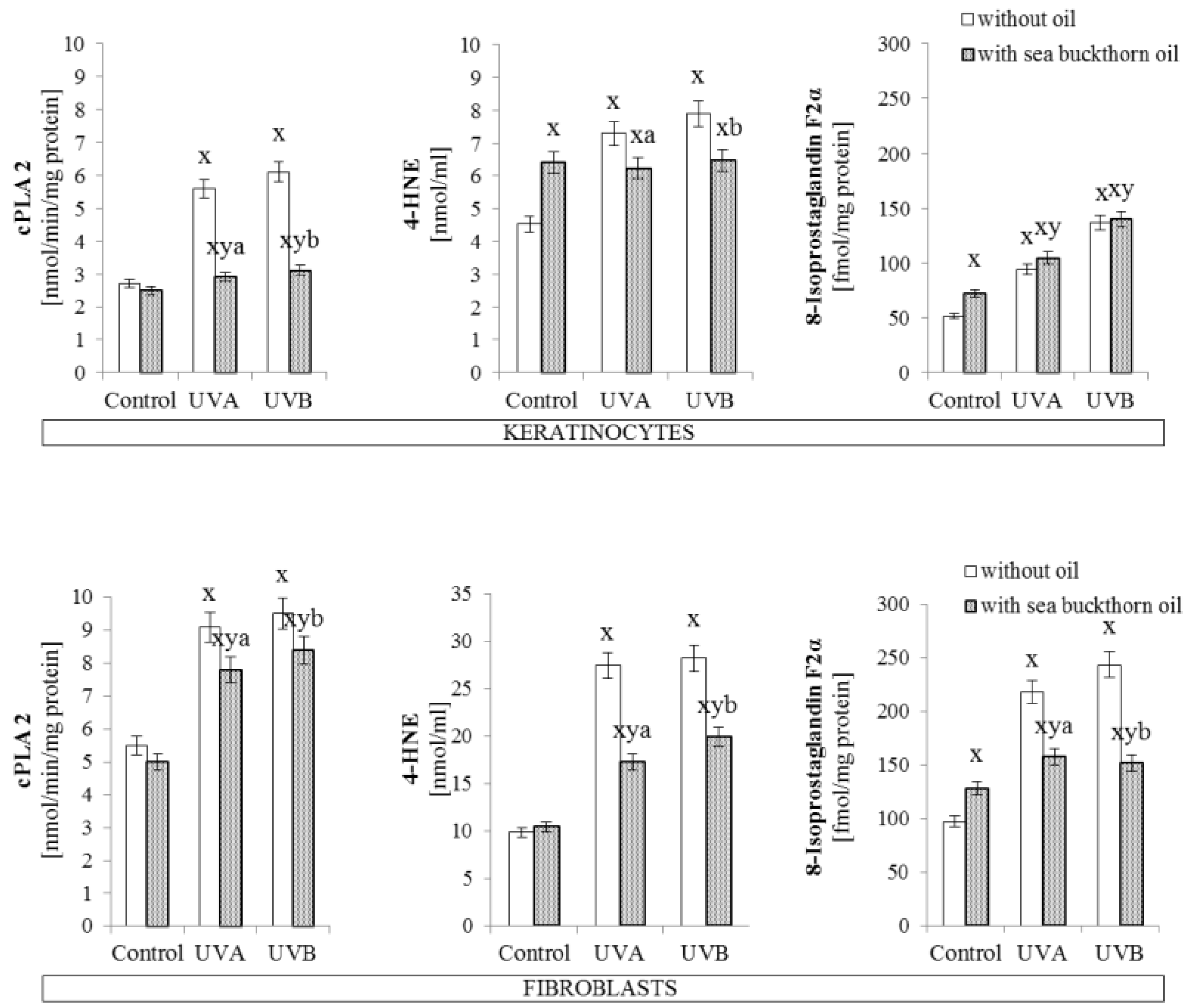
2.5.3. Phospholipase A2 Activity
2.5.4. Determination of Lipid Peroxidation Products
2.5.5. Determination of Endocannabinoid System
2.6. Statistical Analysis
3. Results
3.1. The Sea Buckthorn Seed Oil Composition
3.2. The Effect of Sea Buckthorn Seed Oil on Skin Cells Redox Balance
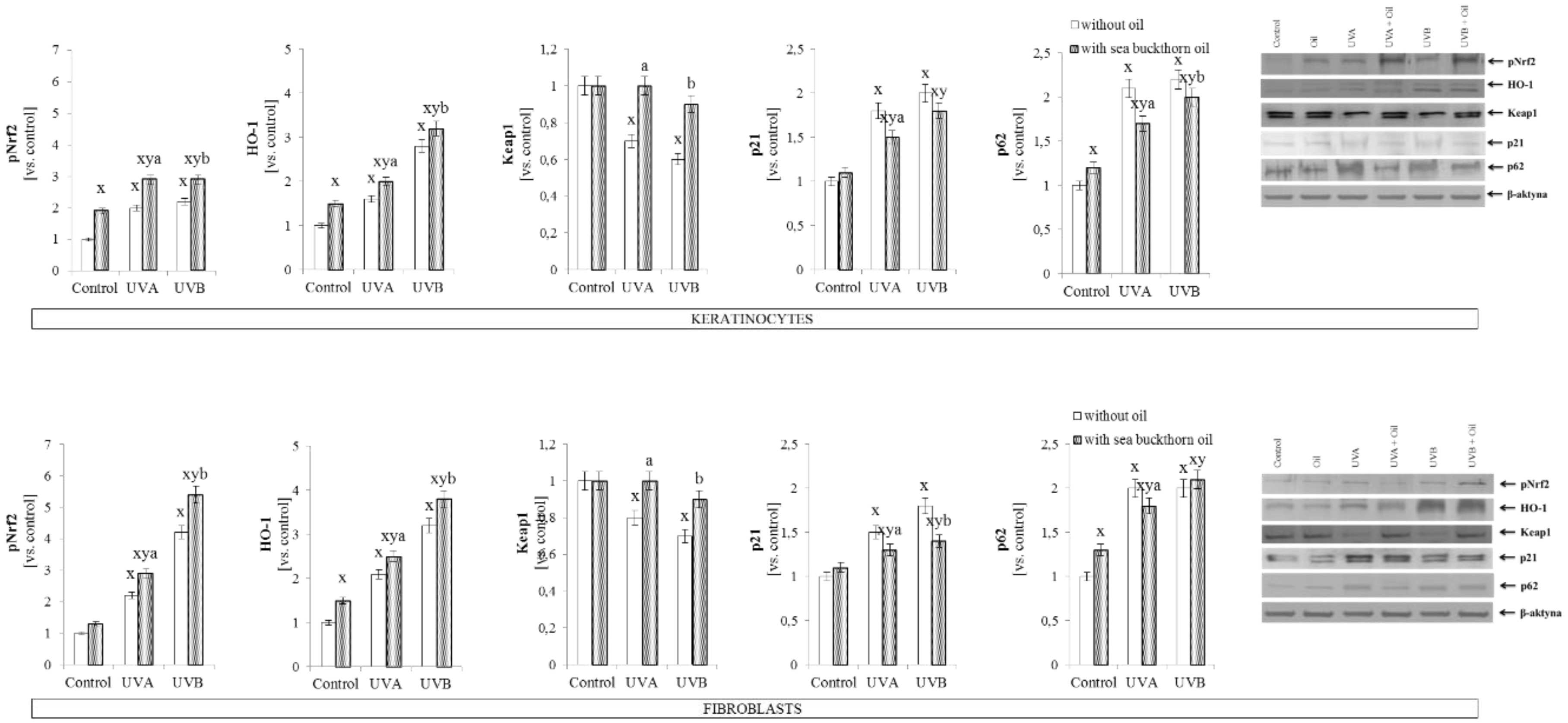
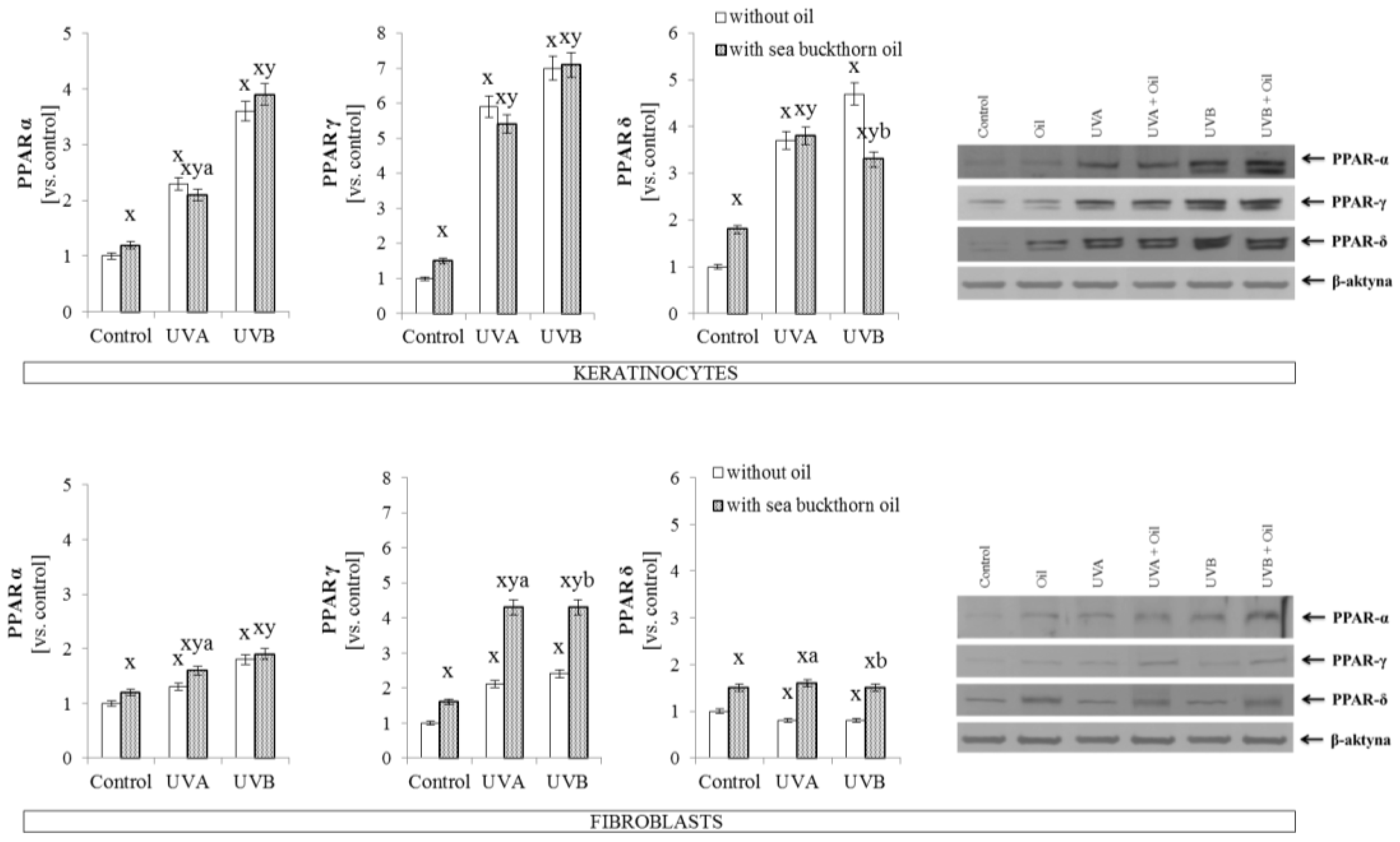
3.3. The Effect of Sea Buckthorn Seed Oil on Skin Cells Transcription Factors
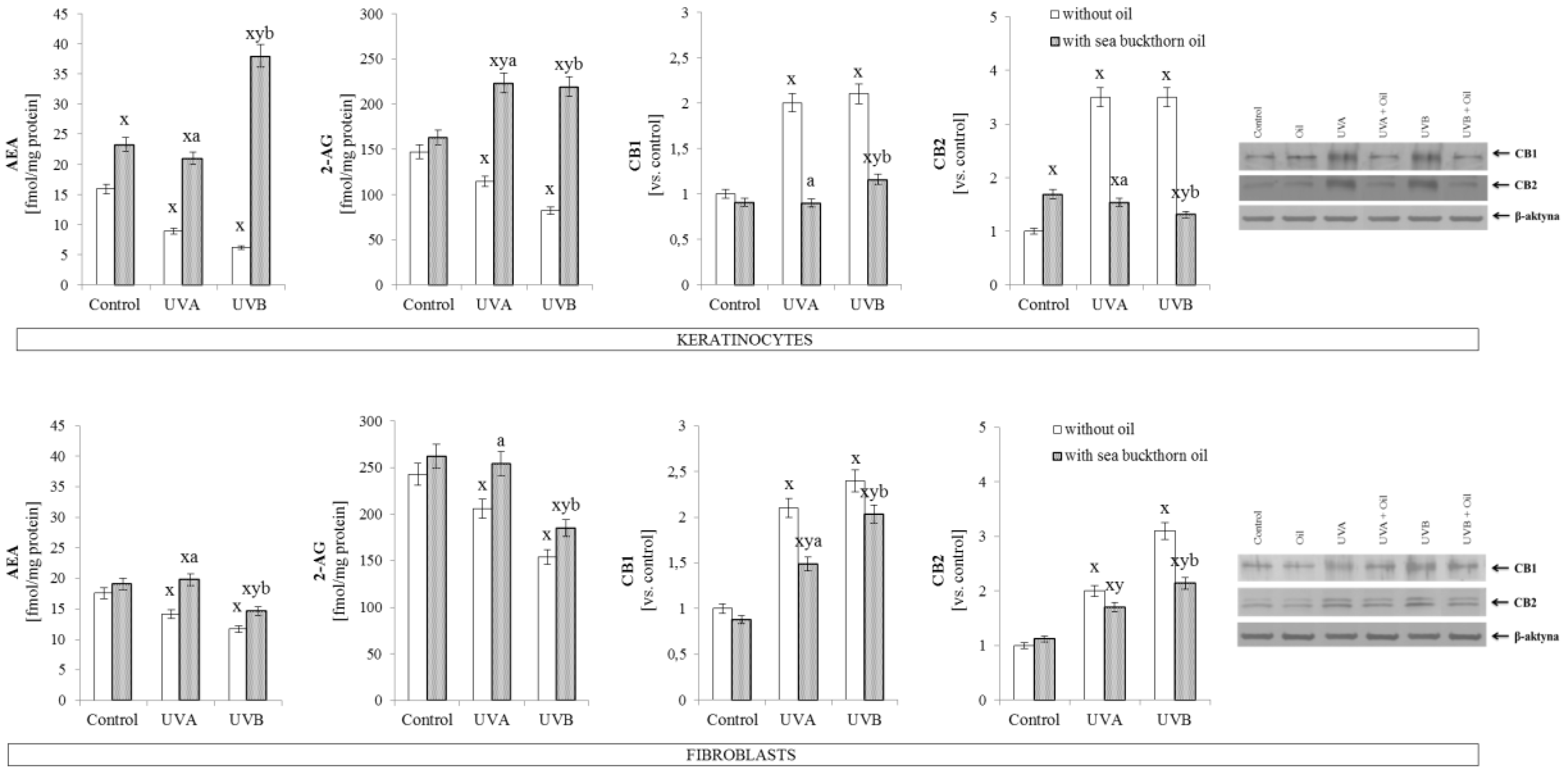
3.4. The Effect of Sea Buckthorn Seed Oil on Skin Cells Lipid Metabolism
4. Discussion
4.1. Sea Buckthorn Seeds Oil Effect on Antioxidant System
4.2. Sea Buckthorn Seeds Oil Effect on Lipid Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chuong, C.M.; Nickoloff, B.J.; Elias, P.M.; Goldsmith, L.A.; Macher, E.; Maderson, P.A.; Sunberg, J.P.; Tagami, H.; Plonka, P.M.; Thestrup-Pederson, K.; et al. What is the ‘ture’ function of skin? Exp. Dermatol. 2002, 11, 159–187. [Google Scholar] [PubMed]
- Agache, P.; Lihoreau, T.; Mac-Mary, S.; Fanian, F.; Humbert, P. The human skin: An overview. In Agache’s Measuring the Skin: Non-invasive Investigations; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 1–4. [Google Scholar]
- Moan, J.; Grigalavicius, M.; Baturaite, Z.; Dahlback, A.; Juzeniene, A. The relationship between UV exposure and incidence of skin cancer. Photodermatol. Photoimmunol. Photomed. 2015, 31, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, V.T.; Ganju, P.; Ramkumar, A.; Grover, R.; Gokhale, R.S. Multifaceted pathways protect human skin from UV radiation. Nat. Chem. Biol. 2014, 10, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV Light Touches the Brain and Endocrine System Through Skin, and Why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Biernacki, M.; Ambrożewicz, E.; Surażyński, A.; Wroński, A.; Skrzydlewska, E. The cross-talk between electrophiles, antioxidant defence and the endocannabinoid system in fibroblasts and keratinocytes after UVA and UVB irradiation. J. Dermatol. Sci. 2016, 81, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.A.A.; Jung, M.; Lee, S.M.; Lee, T.H.; Kim, J. Protective effect of Disporum sessile D. Don extract against UVB-induced photoaging via suppressing MMP-1 expression and collagen degradation in human skin cells. J. Photochem. Photobiol. B 2014, 133, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Larroque-Cardoso, P.; Camaré, C.; Nadal-Wollbold, F.; Grazide, M.H.; Pucelle, M.; Garoby-Salom, S.; Bogdanowicz, P.; Josse, G.; Schmitt, A.M.; Uchida, K.; et al. Elastin modification by 4-hydroxynonenal in hairless mice exposed to UV-A. Role in photoaging and actinic elastosis. J. Investig. Dermatol. 2015, 135, 1873–1881. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Bielawska, K.; Biernacki, M.; Dobrzyńska, I.; Skrzydlewska, E. Time-dependent effect of rutin on skin fibroblasts membrane disruption following UV radiation. Redox Biol. 2017, 12, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Bielawska, K.; Biernacki, M.; Zaręba, I.; Surażyński, A.; Skrzydlewska, E. Comparison of protective effect of ascorbic acid on redox and endocannabinoid systems interactions in in vitro cultured human skin fibroblasts exposed to UV radiation and hydrogen peroxide. Arch. Dermatol. Res. 2017, 309, 285–303. [Google Scholar] [CrossRef] [PubMed]
- Lutz, B.; Marsicano, G.; Maldonado, R.; Hillard, C.J. The endocannabinoid system in guarding against fear, anxiety and stress. Nat. Rev. Neurosci. 2015, 16, 705–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasperi, V.; Dainese, E.; Oddi, S.; Sabatucci, A.; Maccarrone, M. GPR55 and its interaction with membrane lipids: Comparison with other endocannabinoid-binding receptors. Curr. Med. Chem. 2013, 20, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Masaki, H. Role of antioxidants in the skin: Anti-aging effects. J. Dermatol. Sci. 2010, 58, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Kamal-Eldin, A. Effect of fatty acids and tocopherols on the oxidative stability of vegetable oils. Eur. J. Lipid. Sci. Technol. 2006, 108, 1051–1061. [Google Scholar] [CrossRef]
- Suryakumar, G.; Gupta, A. Medicinal and therapeutic potential of Sea buckthorn (Hippophae rhamnoides L.). J. Ethnopharmacol. 2011, 138, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Ting, H.C.; Hsu, Y.W.; Tsai, C.F.; Lu, F.J.; Chou, M.C.; Chen, W.K. The in vitro and in vivo antioxidant properties of seabuckthorn (Hippophae rhamnoides L.) seed oil. Food Chem. 2011, 125, 652–659. [Google Scholar] [CrossRef]
- Zheng, L.; Shi, L.K.; Zhao, C.W.; Jin, Q.Z.; Wang, X.G. Fatty acid, phytochemical, oxidative stability and in vitro antioxidant property of sea buckthorn (Hippophaë rhamnoides L.) oils extracted by supercritical and subcritical technologies. LWT-Food Sci. Technol. 2017, 86, 507–513. [Google Scholar] [CrossRef]
- Teleszko, M.; Wojdyło, A.; Rudzińska, M.; Oszmiański, J.; Golis, T. Analysis of lipophilic and hydrophilic bioactive compounds content in sea buckthorn (Hippophae rhamnoides L.) berries. J. Agric. Food Chem. 2015, 63, 4120–4129. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.M.; Gupta, A.K.; Ahmed, Z.; Kumar, A. Antibacterial and antifungal activity in leaf, seed extract and seed oil of seabuckthorn (Hippophae salicifolia D. Don) plant. J. Plant Pathol. Microbiol. 2011, 2, 1–4. [Google Scholar]
- Punia, D.; Kumari, N. Potential health benefits of Sea buckthorn oil—A review. Agric. Rev. 2017, 38, 233–237. [Google Scholar]
- Hou, D.D.; Di, Z.H.; Qi, R.Q.; Wang, H.X.; Zheng, S.; Hong, Y.X.; Guo, H.; Chen, H.D.; Gao, X.H. Sea Buckthorn (Hippophaë rhamnoides L.) Oil Improves Atopic Dermatitis-Like Skin Lesions via Inhibition of NF-κB and STAT1 Activation. Skin Pharmacol. Physiol. 2017, 30, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Nang Lau, H.L.; Puah, C.W.; Choo, Y.M.; Ma, A.N.; Chuah, C.H. Simultaneous quantification of free fatty acids, free sterols, squalene, and acylglycerol molecular species in palm oil by high-temperature gas chromatography-flame ionization detection. Lipids 2005, 40, 523–528. [Google Scholar] [CrossRef]
- Czaplicki, S.; Ogrodowska, D.; Derewiaka, D.; Tańska, M.; Zadernowski, R. Bioactive compounds in unsaponifiable fraction of oils from unconventional sources. Eur. J. Lipid Sci. Technol. 2011, 113, 1456–1464. [Google Scholar] [CrossRef]
- Zhao, B.; Tham, S.Y.; Lu, J.; Lai, M.H.; Lee, L.K.; Moochhala, S.M. Simultaneous determination of vitamins C, E and β-carotene in human plasma by high-performance liquid chromatography with photodiode-array detection. J. Pharm. Sci. 2004, 7, 200–204. [Google Scholar]
- Fotakis, G.; Timbrell, J.A. In vitro cytotoxicity assays: Comparison of LDH, neutral red, MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride. Toxicol. Lett. 2006, 160, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Kuzkaya, N.; Weissmann, N.; Harrison, D.G.; Dikalov, S. Interactions of peroxynitrite, tetrahydrobiopterin, ascorbic acid, and thiols: Implications for uncoupling endothelial nitricoxide synthase. J. Biol. Chem. 2003, 278, 22546–22554. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Minieri, C.A.; Ollerenshaw, J.D.; Alexander, R.W. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ. Res. 1994, 74, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Prajda, N.; Weber, G. Malignant transformation-linked imbalance: Decreased xanthine oxidase activity in hepatomas. FEBS. Lett. 1975, 59, 245–259. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Mize, C.E.; Longdon, R.G. Hepatic glutathione reductase. Purification and general kinetic properties. J. Biol. Chem. 1962, 237, 1589–1595. [Google Scholar] [PubMed]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [PubMed]
- Sykes, J.A.; McCormac, F.X.; O’Breien, T.J. Preliminary study of the superoxide dismutase content of some human tumors. Cancer Res. 1978, 38, 2759–2762. [Google Scholar] [PubMed]
- Holmgren, A. Thioredoxin and thioredoxin reductase. Methods Enzymol. 1995, 252, 199–208. [Google Scholar] [PubMed]
- Lovell, M.A.; Xie, C.; Gabbita, S.P.; Markesbery, W.R. Decreased thioredoxin and increased thioredoxin reductase levels in Alzheimer’s disease brain. Free Radic. Biol. Med. 2000, 28, 418–427. [Google Scholar] [CrossRef]
- Maeso, N.; Garcia-Martinez, D.; Ruperez, F.J.; Cifuentes, A.; Barbas, C. Capillary electrophoresis of glutathione to monitor oxidative stress and response to antioxidant treatments in an animal model. J. Chromatogr. B 2005, 822, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Vatassery, G.T.; Brin, M.F.; Fahn, S.; Kayden, H.J.; Traber, M.G. Effect of high doses of dietary vitamin E on the concentrations of vitamin E in several brain regions, plasma, liver, and adipose tissue of rats. J. Neurochem. 1988, 51, 621–623. [Google Scholar] [CrossRef] [PubMed]
- Eissa, S.; Seada, L.S. Quantitation of bcl-2 protein in bladder cancer tissue by enzyme immunoassay: Comparison with Western blot and immunohistochemistry. Clin. Chem. 1998, 44, 1423–1429. [Google Scholar] [PubMed]
- Christie, W.W. Preparation of ester derivatives of fatty acids for chromatographic analysis. In Advances in Lipid Methodology; Oily Press: Dundee, UK, 1993; pp. 69–111. [Google Scholar]
- Reynolds, L.J.; Hughes, L.L.; Yu, L.; Dennis, E.A. 1-Hexadecyl-2-arachidonoylthio-2-deoxy-sn-glycero-3-phosphorylcholine as a substrate for the microtiterplate assay of human cytosolic phospholipase A2. Anal. Biochem. 1994, 217, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.P.; Yazdanpanah, M.; Bhooi, N.; Lehotay, D.C. Determination of aldehydes and other lipid peroxidation products in biological samples by gas chromatography-mass spectrometry. Anal. Biochem. 1995, 228, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Coolen, S.A.; van Buuren, B.; Duchateau, G.; Upritchard, J.; Verhagen, H. Kinetics of biomarkers: Biological and technical validity of isoprostanes in plasma. Amino Acids 2005, 29, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Lam, P.M.; Marczylo, T.H.; El-Talatini, M.; Finney, M.; Nallendran, V.; Taylor, A.H.; Konje, J.C. Ultra-performance liquid chromatography tandem mass spectrometry method for the measurement of anandamide in human plasma. Anal. Biochem. 2008, 380, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Bakry, A.M.; Abbas, S.; Ali, B.; Majeed, H.; Abouelwafa, M.Y.; Mousa, A.; Liang, L. Microencapsulation of oils: A comprehensive review of benefits, techniques, and applications. Compr. Rev. Food Sci. Food Saf. 2016, 15, 143–182. [Google Scholar] [CrossRef]
- Gendaszewska-Darmach, E.; Majewska, I. Proangiogenic activity of plant extract in accelerating wound healing—A new face of old phytomedicines. Acta Biochim. Pol. 2011, 58, 449–460. [Google Scholar]
- Yang, B.; Kalimo, K.O.; Tahvonen, R.L.; Mattila, L.M.; Katajisto, J.K.; Kallio, H.P. Effect of dietary supplementation with sea buckthorn (Hippophae rhamnoides) seed and pulp oils on the fatty acid composition of skin glycerophospholipids of patients with atopic dermatitis. J. Nutr. Biochem. 2000, 11, 338–340. [Google Scholar] [CrossRef]
- Zeb, A. Important therapeutic uses of sea buckthorn (Hippophae): A review. J. Biol. Sci. 2004, 4, 687–693. [Google Scholar]
- Michel, T.; Destandau, E.; Le Flochc, G.; Lucchesi, M.E.; Elfakir, C. Antimicrobial, antioxidant and phytochemical investigations of sea buckthorn (Hippophae rhamnoides L.) leaf, stem, root and seed. Food Chem. 2012, 131, 754–760. [Google Scholar] [CrossRef]
- Kumar, M.S.Y.; Dutta, R.; Prasad, D.; Mishra, K. Subcritical water extraction of antioxidant compounds from Seabuckthorn (Hippophae rhamnoides L.) leaves for the comparative evaluation of antioxidant activity. Food Chem. 2011, 127, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Baccarin, T.; Mitjans, M.; Ramos, D.; Lemos-Senna, E.; Vinardell, M.P. Photoprotection by Punica granatum seed oil nanoemulsion entrapping polyphenol-rich ethyl acetate fraction against UVB-induced DNA damage in human keratinocyte (HaCaT) cell line. J. Photochem. Photobiol. B 2015, 153, 127–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manku, M.S.; Horrobin, D.F.; Morse, N.L.; Wright, S.; Burton, J.L. Essential fatty acids in the plasma phospholipids of patients with atopic eczema. Br. J. Dermatol. 1984, 110, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Ganju, L.; Padwad, Y.; Singh, R.; Karan, D.; Chanda, S.; Chopra, M.K.; Bhatnagar, P.; Kashyap, R.; Sawhney, R.C. Anti-inflammatory activity of Seabuckthorn (Hippophae rhamnoides) leaves. Int. Immunopharmacol. 2005, 5, 1675–1684. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S.Y.; Maheshwari, D.T.; Verma, S.K.; Singh, V.K.; Singh, S.N. Antioxidant and hepatoprotectice activities of phenolic rich fraction of Seabuckthorn (Hippophae rhamnoides L.) leaves. Food Chem. Toxicol. 2011, 49, 2422–2428. [Google Scholar]
- Myong-Jo, K.; Ju-Sung, K.; Yong-Soo, K.; Yeo-Jin, S. Isolation and identification of Sea buckthorn (Hippophae rhamnoides) phenolics with antioxidant activity and α-glucosidase inhibitory effect. J. Agric. Food Chem. 2011, 59, 138–144. [Google Scholar]
- Schallreuter, K.U.; Wood, J.M. The role of thioredoxin reductase in the reduction of free radicals at the surface of the epidermis. Biochem. Biophys. Res. Commun. 1986, 136, 630–637. [Google Scholar] [CrossRef]
- Arner, E.S.; Holmgren, A. The thioredoxin system in cancer. In Seminars in Cancer Biology; Academic Press: Orlando, FL, USA, 2006. [Google Scholar]
- Carvalho, C.M.; Chew, E.H.; Hashemy, S.I.; Lu, J.; Holmgren, A. Inhibition of the human thioredoxin system a molecular mechanism of mercury toxicity. J. Biol. Chem. 2008, 283, 11913–11923. [Google Scholar] [CrossRef] [PubMed]
- Sougrakpam, Y.; Deswal, R. Hippophae rhamnoides N-glycoproteome analysis: A small step towards sea buckthorn proteome mining. Physiol. Mol. Biol. Plants 2016, 22, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Rybałtowska-Kawałko, P.; Skrzydlewska, E. Rutin as a mediator of lipid metabolism and cellular signaling pathways interactions in fibroblasts altered by UVA and UVB radiation. Oxid Med. Cell Longev. 2017, 2017, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Skrzydlewska, E. The role of transcription factor Nrf2 in skin cells metabolism. Arch. Dermatol. Res. 2015, 307, 385–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konstantinopoulos, P.A.; Spentzos, D.; Fountzilas, E.; Francoeur, N.; Sanisetty, S.; Grammatikos, A.P.; Hecht, J.L.; Cannistra, S.A. Keap1 mutations and Nrf2 pathway activation in epithelial ovarian cancer. Cancer Res. 2011, 71, 5081–5089. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, W.; Kong, A.N. Anti-oxidative stress regulator NF-E2-related factor 2 mediates the adaptive induction of antioxidant and detoxifying enzymes by lipid peroxidation metabolite 4-hydroxynonenal. Cell Biosci. 2012, 2, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes. Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abiko, Y.; Kumagai, Y. Interaction of Keap1 modified by 2-tert-butyl-1, 4-benzoquinone with GSH: Evidence for S-transarylation. Chem. Res. Toxicol. 2013, 26, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.; Wang, H.; Xu, J.; Li, T.; Zhang, L.; Ding, Y.; Zhu, L.; He, J.; Zhou, M.L. Melatonin stimulates antioxidant enzymes and reduces oxidative stress in experimental traumatic brain injury: The Nrf2–ARE signaling pathway as a potential mechanism. Free Radic. Biol. Med. 2014, 73, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.W.; Sweatman, T.W.; Semak, I.; Sayre, R.M.; Wortsman, J.; Slominski, A. Constitutive and UV-induced metabolism of melatonin in keratinocytes and cell-free systems. FASEB J. 2006, 20, 1564–1566. [Google Scholar] [CrossRef] [PubMed]
- Janjetovic, Z.; Jarrett, S.G.; Lee, E.F.; Duprey, C.; Reiter, R.J.; Slominski, A.T. Melatonin and its metabolites protect human melanocytes against UVB-induced damage: Involvement of NRF2-mediated pathways. Sci. Rep. 2014, 7, 1274. [Google Scholar] [CrossRef] [PubMed]
- Ranjith, A.; Kumar, K.S.; Venugopalan, V.V.; Arumughan, C.; Sawhney, R.C.; Singh, V. Fatty acids, tocols, and carotenoids in pulp oil of three sea buckthorn species (Hippophae rhamnoides, H. salicifolia, and H. tibetana) grown in the Indian Himalayas. J. Am. Oil Chem. Soc. 2006, 83, 359–364. [Google Scholar] [CrossRef]
- Erkkola, R.; Yang, B. Sea buckthorn oils: Towards healthy mucous membranes. Agro Food Ind. Hi Tech 2003, 14, 53–59. [Google Scholar]
- Piłat, B.; Zadernowski, R. Rokitnik w produktach spożywczych. Przemysł Spożywczy 2016, 70, 35–38. [Google Scholar] [CrossRef]
- Welters, H.J.; Diakogiannaki, E.; Mordue, J.M.; Tadayyon, M.; Smith, S.A.; Morgan, N.G. Differential protective effects of palmitoleic acid and cAMP on caspase activation and cell viability in pancreatic β-cells exposed to palmitate. Apoptosis 2006, 11, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, R.; Hildebrand, D.F. Biosynthesis and metabolic engineering of palmitoleate production, an important contributor to human health and sustainable industry. Prog. Lipid Res. 2012, 51, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H.; Miyahara, H.; Hatanaka, A. Chronic administration of palmitoleic acid reduces insulin resistance and hepatic lipid accumulation in KK-A y Mice with genetic type 2 diabetes. Lipids Health Dis. 2011, 10, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Guo, Q.; Qin, C.; Shang, R.; Zhang, Z. Sea buckthorn fruit oil extract alleviates insulin resistance through the PI3K/Akt signaling pathway in type 2 diabetes mellitus cells and rats. J. Agric. Food Chem. 2017, 65, 1328–1336. [Google Scholar] [CrossRef] [PubMed]
- Bolsoni-Lopes, A.; Festuccia, W.T.; Chimin, P.; Farias, T.S.; Torres-Leal, F.L.; Cruz, M.M.; Andrade, P.B.; Hirabara, S.M.; Lima, F.B.; Alonso-Vale, M.I.C. Palmitoleic acid (n-7) increases white adipocytes GLUT4 content and glucose uptake in association with AMPK activation. Lipids Health Dis. 2014, 13, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Białek, M.; Rutkowska, J. The importance of γ-linolenic acid in the prevention and treatment. Adv. Hyg. Exp. Med. 2015, 69, 892–904. [Google Scholar] [CrossRef]
- Larmo, P.S.; Järvinen, R.L.; Setälä, N.L.; Yang, B.; Viitanen, M.H.; Engblom, J.R.K.; Tahvonen, R.L.; Kallio, H.P. Oral Sea Buckthorn Oil Attenuates Tear Film Osmolarity and Symptoms in Individuals with Dry Eye–4. J. Nutr. 2010, 140, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Sergeant, S.; Rahbar, E.; Chilton, F.H. Gamma-linolenic acid, Dihommo-gamma linolenic, eicosanoids and inflammatory processes. Eur. J. Pharmacol. 2016, 785, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Luczaj, W.; Gęgotek, A.; Skrzydlewska, E. Antioxidants and HNE in redox homeostasis. Free Radic. Biol. Med. 2017, 111, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Baranowska-Bosiacka, I.; Gutowska, I.; Chlubek, D. The effect of reactive oxygen species on the synthesis of prostanoids from arachidonic acid. J. Physiol. Pharmacol. 2013, 64, 409–421. [Google Scholar] [PubMed]
- Davis, M.P. Cannabinoids in pain management: CB1, CB2 and non-classic receptor ligands. Expert Opin. Investig. Drugs 2014, 23, 1123–1140. [Google Scholar] [CrossRef] [PubMed]
- Gresham, A.; Masferrer, J.; Chen, X.; Leal-Khouri, S.; Pentland, A.P. Increased synthesis of high-molecular-weight cPLA2 mediates early UV-induced PGE2 in human skin. Am. J. Physiol. 1996, 270, C1037–C1050. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Bae, S.K.; Pyo, M.; Heo, Y.; Kim, C.G.; Kang, C.; Kim, E. Anti-wrinkle effect of PLA2-free bee venom against UVB-irradiated human skin cells. J. Agric. Life Sci. 2015, 49, 125–135. [Google Scholar] [CrossRef]
- Montserrat-De, L.P.S.; García-Giménez, M.D.; Ángel-Martín, M.; Pérez-Camino, M.C.; Arche, A.F. Long-chain fatty alcohols from evening primrose oil inhibit the inflammatory response in murine peritoneal macrophages. J. Ethnopharmacol. 2014, 151, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Kalimo, K.O.; Mattila, L.M.; Kallio, S.E.; Katajisto, J.K.; Peltola, O.J.; Kallio, H.P. Effects of dietary supplementation with sea buckthorn (Hippophae rhamnoides) seed and pulp oils on atopic dermatitis. J. Nutr. Biochem. 1999, 10, 622–630. [Google Scholar] [CrossRef]
- Paus, R. The Skin and endocrine disorders. In Rook’s Textbook of Dermatology, 9th ed.; Wiley Online Library: Hoboken, NJ, USA, 2016; pp. 1–30. [Google Scholar]
- Skobowiat, C.; Postlethwaite, A.E.; Slominski, A.T. Skin exposure to ultraviolet B rapidly activates systemic neuroendocrine and immunosuppressive responses. Photochem. Photobiol. 2017, 93, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.D.; Prabhu, K.S.; Thompson, J.T.; Reddy, P.S.; Peters, J.M.; Peterson, B.R.; Reddy, C.C.; Heuvel, J.P.V. The oxidative stress mediator 4-hydroxynonenal is an intracellular agonist of the nuclear receptor peroxisome proliferator-activated receptor-β/δ (PPARβ/δ). Free Radic. Biol. Med. 2007, 42, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Wahli, W.; Michalik, L. PPARs at the crossroads of lipid signaling and inflammation. Trends Endocrinol. Metab. 2012, 23, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Subramaniyan, S.A.; Kim, S.; Hwang, I. Cell-Cell Communication between fibroblast and 3T3-L1 cells under co-culturing in oxidative stress condition induced by H2O2. Appl. Biochem. Biotechnol. 2016, 180, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Benito, C.; Tolón, R.M.; Castillo, A.I.; Ruiz-Valdepeñas, L.; Martínez-Orgado, J.A.; Fernández-Sánchez, F.J.; Vázquez, C.; Cravatt, B.F.; Romero, J. β−Amyloid exacerbates inflammation in astrocytes lacking fatty acid amide hydrolase through a mechanism involving PPAR-α, PPAR-γ and TRPV1, but not CB1 or CB2 receptors. Br. J. Pharmacol. 2012, 166, 1474–1489. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Hsu, W.H.; Liao, T.H.; Pan, T.M. The monascus metabolite monascin against TNF-α-induced insulin resistance via suppressing PPAR-γ phosphorylation in C2C12 myotubes. Food Chem. Toxicol. 2011, 49, 2609–2617. [Google Scholar] [CrossRef] [PubMed]
- Pichiah, P.T.; Moon, H.J.; Park, J.E.; Moon, Y.J.; Cha, Y.S. Ethanolic extract of seabuckthorn (Hippophae rhamnoides L.) prevents high-fat diet–Induced obesity in mice through down-regulation of adipogenic and lipogenic gene expression. Nutr. Res. 2012, 32, 856–864. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids Composition of Sea Buckthorn Needs Oil | |||||
| PL | TG | DG | FFA | ||
| mg/mL | |||||
| 14:0 | 0.060 ± 0.003 | 4.58 ± 0.14 | 0.240 ± 0.007 | 0.070 ± 0.003 | |
| 16:0 | 1.14 ± 0.03 | 308.60 ± 9.25 | 3.10 ± 0.09 | 1.65 ± 0.05 | |
| 18:0 | 0.51 ± 0.02 | 8.05 ± 0.24 | 0.47 ± 0.01 | 0.100 ± 0.003 | |
| 20:0 | n.d. | 2.13 ± 0.06 | 0.130 ± 0.003 | 0.060 ± 0.002 | |
| 22:0 | n.d. | 0.95 ± 0.03 | 0.140 ± 0.003 | n.d. | |
| 24:0 | n.d. | 1.08 ± 0.03 | 0.170 ± 0.004 | 0.070 ± 0.002 | |
| SFA | 1.71 ± 0.05 | 325.40 ± 9.76 | 4.26 ± 0.13 | 1.95 ± 0.06 | |
| 16:1n7 | 0.48 ± 0.01 | 310.85 ± 9.33 | 4.56 ± 0.14 | 2.22 ± 0.07 | |
| 18:1n9c | 0.150 ± 0.005 | 45.34 ± 1.36 | 0.89 ± 0.03 | 0.34 ± 0.01 | |
| 18:1n7 | 0.110 ± 0.003 | 45.25 ± 1.35 | 0.83 ± 0.02 | 0.48 ± 0.01 | |
| 18:2n6 | 0.230 ± 0.007 | 87.68 ± 2.63 | 7.15 ± 0.21 | 1.53 ± 0.05 | |
| 18:3γ | n.d. | 1.79 ± 0.05 | n.d. | n.d. | |
| 18:3α | 0.280 ± 0.008 | 7.05 ± 0.21 | n.d. | 0.210 ± 0.006 | |
| MUFA | 0.74 ± 0.02 | 401.44 ± 12.04 | 6.29 ± 0.19 | 3.04 ± 0.09 | |
| PUFA | 0.51 ± 0.02 | 96.51 ± 2.90 | 7.15 ± 0.21 | 1.74 ± 0.05 | |
| USFA | 1.25 ± 0.04 | 497.95 ± 14.94 | 13.44 ± 0.40 | 4.78 ± 0.14 | |
| Sum | 2.95 ± 0.09 | 823.35 ± 24.70 | 17.70 ± 0.53 | 6.73 ± 0.20 | |
| Antioxidant Components of Sea Buckthorn Needs Oil [mg/100 g] | |||||
| Squalene | β-Carotene | Vitamin E | Vitamin A | ||
| 240±11 | 15.11 ± 0.46 | 98.92 ± 2.89 | 648.0 ± 19.7 | ||
| Phytosterols in Sea Buckthorn Needs Oil [mg/100 g] | |||||
| Cholesterol | Brassicasterol | Campesterol | Stigmasterol | β-Sitosterol | Total phytosterols |
| 0.51 ± 0.01 | 4.90 ± 0.14 | 17.61 ± 0.48 | 4.95 ± 0.12 | 223.88 ± 6.76 | 251.87 ± 7.48 |
| Prooxidative Parameters | Keratinocytes | Fibroblasts | |||||
|---|---|---|---|---|---|---|---|
| Oil | Control | UVA | UVB | Control | UVA | UVB | |
| ROS [nM/min/mg protein] | - | 32.8 ± 1.6 | 89.1 ± 4.3 x | 98.9 ± 4.8 x | 50.6 ±2.4 | 126.5 ± 6.1 x | 132.1 ± 6.4 x |
| + | 63.5 ± 3.1 x | 80.8 ± 3.9 xy | 96.5 ± 4.7 xy | 54.7 ±2.6 | 94.6 ± 4.6 xya | 96.5 ± 4.7 xyb | |
| NOX [RLU/mg protein] | - | 158 ± 7.7 | 229 ± 11.2 x | 256 ± 12.5 x | 179 ± 8.7 | 321 ± 15.7 x | 398 ± 19.5 x |
| + | 173 ± 8.4 | 231 ± 11.3 xy | 227 ± 14.0 xyb | 216 ± 10.5 x | 268 ± 13.1 xya | 321 ± 15.7 xyb | |
| XO [mU/mg protein] | - | 164 ± 8.1 | 267 ± 13.1 x | 297 ± 14.5 x | 114 ± 5.5 | 248 ± 12.1 x | 487 ± 23.8 x |
| + | 158 ± 7.7 | 248 ± 12.1 xy | 254 ± 12.4 xyb | 278 ± 13.6 x | 215 ± 10.5 xya | 348 ± 17.1 xyb | |
| Antioxidant Parameters | Keratinocytes | Fibroblasts | |||||
|---|---|---|---|---|---|---|---|
| Oil | Control | UVA | UVB | Control | UVA | UVB | |
| GSH-Px [mU/mg protein] | - | 13.6 ± 0.6 | 10.8 ± 0.5 x | 7.8 ± 0.4 x | 11.8 ± 0.6 | 26.4 ± 1.2 x | 29.5 ± 1.4 x |
| + | 15.6 ± 0.7 x | 12.4 ± 0.6 ya | 14.8 ± 0.7 b | 16.3 ± 0.8 x | 26.9 ± 1.3 xy | 31.2 ± 1.5 xy | |
| GSSG-R [mU/mg protein] | - | 26.3 ± 1.2 | 23.8 ± 1.2 x | 19.2 ± 0.9 x | 22.3 ± 1.1 | 45.6 ± 2.2 x | 56.2 ± 2.7 x |
| + | 22.5 ± 1.1 x | 21.8 ± 1.1 x | 18.7 ± 0.9 xy | 26.9 ± 1.3 x | 34.6 ± 1.7 xy | 49.3 ± 2.4 xy | |
| SOD [mU/mg protein] | - | 28.4 ± 1.3 | 23.1 ± 1.1 x | 21.6 ± 1.1 x | 26.3 ± 1.2 | 21.3 ± 1.0 x | 14.6 ± 0.7 x |
| + | 30.5 ± 1.5 | 25.6 ± 1.2 xy | 24.7 ± 1.2 xyb | 20.6 ± 1.0 x | 18.5 ± 0.9 xya | 12.3 ± 0.6 xyb | |
| TrxR [µU/mg protein] | - | 112 ± 5 | 93 ± 4 x | 84 ± 4 x | 331 ± 16 | 204 ± 10 x | 141 ± 7 x |
| + | 104 ± 5 | 421 ± 11 xya | 300 ± 9 xyb | 312 ± 15 | 243 ± 11 xya | 182 ± 8 xyb | |
| Trx [µg/mg protein] | - | 1.5 ± 0.07 | 1 ± 0.07 x | 0.8 ± 0.07 x | 0.9 ± 0.04 | 0.4 ± 0.01 x | 0.4 ± 0.02 x |
| + | 1.6 ± 0.08 | 1.6 ± 0.08 xa | 1.1 ± 0.07 xb | 1.5 ± 0.07 x | 1 ± 0.04 xya | 1.1 ± 0.05 xyb | |
| GSH [nmol/mg protein] | - | 26.2 ± 1.2 | 11.4 ± 0.5 x | 9.6 ± 0.4 x | 14.6 ± 0.7 | 8.2 ± 0.4 x | 5.6 ± 0.3 x |
| + | 37.7 ± 1.8 x | 27.7 ± 1.3 ya | 22.3 ± 1.1 xyb | 15.6 ± 0.8 | 10.4 ± 0.5 xya | 10.1 ± 0.5 xyb | |
| vitamin A [nmol/mg protein] | - | 53 ± 2.5 | 39 ± 1.9 x | 35 ± 1.7 x | 41 ± 2.1 | 34 ± 1.6 x | 33 ± 1.6 x |
| + | 63 ± 3.0 x | 59 ± 2.8 xa | 56 ± 2.7 yb | 46 ± 2.2 | 41 ± 2.1 a | 38 ± 1.8 xyb | |
| vitamin E [nmol/mg protein] | - | 453 ± 22.1 | 325 ± 15.9 x | 303 ± 14.8 x | 336 ± 16.5 | 287 ±1 4.1 x | 269 ± 13.1 x |
| + | 517 ± 25.3 x | 491 ± 24.1 ya | 478 ± 23.4 yb | 401 ± 19.5 x | 381 ± 18.6 xa | 351 ± 17.2 yb | |
| Keratinocytes | Fibroblasts | ||||||
|---|---|---|---|---|---|---|---|
| Oil | Control | UVA | UVB | Control | UVA | UVB | |
| Phospholipid fatty acids [ug/mg protein] | |||||||
| 14:0 | - | 5.3 ± 0.3 | 4.5 ± 0.2 | 4.5 ± 0.2 | 1.4 ± 0.1 | 1.2 ± 0.1 | 1.2 ± 0.1 |
| + | 12.4 ± 0.6 x | 11.3 ± 0.6 xa | 11.8 ± 0.6 xb | 6.3 ± 0.1 x | 2.5 ± 0.1 xya | 5.1 ± 0.3 xyb | |
| 16:0 | - | 152 ± 7.3 | 129 ± 6.5 x | 128 ± 6.4 x | 80 ± 4 | 68 ± 3.4 | 68 ± 3.4 |
| + | 662 ± 33.1 x | 145 ± 7.3 y | 260 ± 13.0 xyb | 190 ± 9.5 x | 90 ± 4.5 ya | 122 ± 6.1 xyb | |
| 16:1 | - | 15.6 ± 0.8 | 13.2 ± 0.7 | 13.1 ± 0.7 | 13.6 ± 0.7 | 11.6 ± 0.6 | 11.5 ± 0.6 |
| + | 29.9 ± 1.5 x | 27.5 ± 1.4 xa | 28.5 ± 1.4 xb | 23.2 ± 1.2 x | 17.7 ± 0.9 xya | 15.9 ± 0.8 xyb | |
| 18:0 | - | 123 ± 6.2 | 104 ± 5.2 x | 103 ± 5.2 x | 69 ± 3.5 | 58 ± 2.9 x | 58 ± 2.9 x |
| + | 412 ± 20.6 x | 137 ± 6.9 xya | 187 ± 9.4 xyb | 126 ± 6.3 x | 109 ± 5.5 xya | 110 ± 5.5 xyb | |
| 18:1nc | - | 199 ± 10.1 | 169 ± 8.5 x | 168 ± 8.4 x | 112 ± 5.6 | 95 ± 4.8 x | 94 ± 4.7 x |
| + | 580 ± 29.1 x | 129 ± 6.5 xya | 386 ± 19.3 xyb | 181 ± 9.1 x | 167 ± 8.4 xa | 114 ± 5.7 yb | |
| 18:1nt | - | 47 ± 2.4 | 39 ± 2.1 x | 39 ± 2.1 x | 18 ± 0.9 | 16 ± 0.8 | 15 ± 0.8 x |
| + | 170 ± 18.5 x | 48 ± 2.4 ya | 139 ± 7.1 xyb | 58 ± 2.9 x | 22 ± 1.1 xya | 36 ± 1.8 xyb | |
| 18:2 | - | 116.2 ± 5.8 | 98.8 ± 4.9 x | 98.1 ± 4.9 x | 45.4 ± 2.3 | 38.6 ± 1.9 x | 38.3 ± 1.9 x |
| + | 162.7 ± 8.1 x | 111.4 ± 5.6 ya | 137.2 ± 6.9 xyb | 69.8 ± 3.5 x | 60.4 ± 3.0 xya | 64.5 ± 3.2 xb | |
| 18:3n3 | - | 24.6 ± 1.2 | 20.9 ± 1.1 x | 20.8 ± 1.1 x | 6.2 ± 0.3 | 5.2 ± 0.3 | 5.5 ± 0.3 |
| + | 29.5 ± 1.5 x | 30.2 ± 1.5 xa | 31.2 ± 1.6 xb | 12.5 ± 0.6 x | 7.8 ± 0.4 xya | 8.5 ± 0.4 xyb | |
| 20:4 | - | 38.1 ± 1.9 | 32.4 ± 1.6 x | 32.2 ± 1.6 x | 35.6 ± 1.8 | 30.2 ± 1.5 | 30.1 ± 1.5 |
| + | 197.4 ± 9.9 x | 36.4 ± 1.8 ya | 104.7 ± 5.2 xyb | 81.7 ± 4.1 x | 30.3 ± 1.5 y | 57.5 ± 2.9 xyb | |
| 22:6 | - | 11.1 ± 0.6 | 9.4 ± 0.5 | 9.3 ± 0.5 | 8.2 ± 0.4 | 6.9 ± 0.3 | 6.9 ± 0.3 |
| + | 30.1 ± 1.5 x | 18.1 ± 0.9 xya | 23.1 ± 1.2 xyb | 41.6 ± 2.1 x | 16.2 ± 0.8 xya | 33.8 ± 1.7 xyb | |
| Free fatty acids [µg/mg protein] | |||||||
| 16:0 | - | 8.8 ± 0.4 | 7.5 ± 0.4 | 7.4 ± 0.4 | 2.9 ± 0.1 | 2.4 ± 0.1 | 2.4 ± 0.1 |
| + | 26 ± 1.3 x | 14.1 ± 0.7 xya | 19.8 ± 1 xyb | 9.8 ± 0.5 x | 3.9 ± 0.2 xya | 7 ± 0.4 xyb | |
| 16:1 | - | 1.7 ± 0.1 | 1.4 ± 0.1 | 1.4 ± 0.1 | 0.7 ± 0.1 | 0.6 ± 0.1 | 0.6 ± 0.1 |
| + | 5.5 ± 0.3 x | 1.2 ± 0.1 y | 4.4 ± 0.2 xyb | 2.1 ± 0.1 x | 1.3 ± 0.1 xya | 1.6 ± 0.1 xyb | |
| 18:0 | - | 7.6 ± 0.4 | 6.4 ± 0.3 | 6.4 ± 0.3 | 3.4 ± 0.2 | 2.9 ± 0.1 | 2.9 ± 0.3 |
| + | 16.8 ± 0.8 x | 8.8 ± 0.4 ya | 8.2 ± 0.4 yb | 8.8 ± 0.4 x | 3.2 ± 0.2 y | 5.5 ± 0.3 xyb | |
| 18:1nc | - | 13.3 ± 0.7 | 11.3 ± 0.6 | 11.2 ± 0.6 | 3.8 ± 0.2 | 3.3 ± 0.2 | 3.5 ± 0.2 |
| + | 54.3 ± 2.7 x | 51.1 ± 2.6 xa | 45.1 ± 2.3 xyb | 6.8 ± 0.3 x | 4.1 ± 0.2 ya | 5.5 ± 0.3 xyb | |
| 18:1nt | - | 6.1 ± 0.3 | 5.2 ± 0.3 | 5.1 ± 0.3 | 1.1 ± 0.1 | 0.9 ± 0.1 | 0.9 ± 0.1 |
| + | 9.8 ± 0.5 x | 8.5 ± 0.4 xya | 8.3 ± 0.4 xyb | 4.5 ± 0.2 x | 1.5 ± 0.1 xya | 4.1 ± 0.2 xb | |
| 18:2 | - | 11.7 ± 0.6 | 9.9 ± 0.5 | 9.8 ± 0.5 | 3.2 ± 0.2 | 2.7 ± 0.1 | 2.7 ± 0.1 |
| + | 15.1 ± 0.8 x | 10.5 ± 0.5 y | 12.7 ± 0.6 xyb | 5.4 ± 0.3 x | 3.3 ± 0.2 ya | 3.3 ± 0.2 y | |
| 20:4 | - | 2.2 ± 0.1 | 1.8 ± 0.1 | 1.8 ± 0.1 | 0.7 ± 0.1 | 0.6 ± 0.1 | 0.6 ± 0.1 |
| + | 9 ± 0.5 x | 5.4 ± 0.3 xya | 5.6 ± 0.3 xyb | 3.4 ± 0.2 x | 0.6 ± 0.1 y | 2.7 ± 0.1 xyb | |
| 22:6 | - | 0.9 ± 0.1 | 0.8 ± 0.1 | 0.8 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 |
| + | 11.3 ± 0.6 x | 11.4 ± 0.6 xa | 4.8 ± 0.2 xyb | 2.3 ± 0.1 x | 2.2 ± 0.1 xa | 2.2 ± 0.1 xb | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gęgotek, A.; Jastrząb, A.; Jarocka-Karpowicz, I.; Muszyńska, M.; Skrzydlewska, E. The Effect of Sea Buckthorn (Hippophae rhamnoides L.) Seed Oil on UV-Induced Changes in Lipid Metabolism of Human Skin Cells. Antioxidants 2018, 7, 110. https://doi.org/10.3390/antiox7090110
Gęgotek A, Jastrząb A, Jarocka-Karpowicz I, Muszyńska M, Skrzydlewska E. The Effect of Sea Buckthorn (Hippophae rhamnoides L.) Seed Oil on UV-Induced Changes in Lipid Metabolism of Human Skin Cells. Antioxidants. 2018; 7(9):110. https://doi.org/10.3390/antiox7090110
Chicago/Turabian StyleGęgotek, Agnieszka, Anna Jastrząb, Iwona Jarocka-Karpowicz, Marta Muszyńska, and Elżbieta Skrzydlewska. 2018. "The Effect of Sea Buckthorn (Hippophae rhamnoides L.) Seed Oil on UV-Induced Changes in Lipid Metabolism of Human Skin Cells" Antioxidants 7, no. 9: 110. https://doi.org/10.3390/antiox7090110